Chapter 21 Landscape Ecology
Thirteen centuries ago, an emperor of
Countless are the
mountains in Yamato,
But perfect is
the heavenly hill of Kagu;
When I climb it
and survey my realm,
Over the wide lake
the gulls are on the wing;
A beautiful land
it is, the

FIGURE 21.1
The views from hills and mountains, such as this one, were used traditionally
in ancient
Tadahiko Higuchi (i983) cited this poem in his book
on the visual and spatial structure of landscapes. He wondered about
the significance of the emperor's trip to the top of
Long before the emperor Jomei
stood upon
What were these emperors, community leaders,
and farmers doing on their promontories each spring? What were they
looking at? There are many possible reasons for the custom, including
religious, esthetic, economic, and political reasons. Regardless of
the precise origin and motivation for the ritual, the custom of "seeing
the domain" was a source of information about the landscape upon
which these people depended, information of potential value to emperor,
community leader, or individual farmer alike.
Imagine yourself standing on the top of a
mountain as Japanese emperors once did. What would you see on the surrounding
plains? You would see villages, fields, paths, roads, streams, woods,
and much more. What would you do with this information? From these heights
you could see if fields were well kept, if seedling density was uniform
across recently planted fields, if villages were well maintained, if
forests were encroaching on cleared properties, and so forth. Ecologists
define landscape ecology as the study of landscape structure and processes.
Given this definition, kunimi, or domain viewing, may be the first written record
of the practice of landscape ecology.
Though landscape ecology is young as a scientific
discipline, people seem to have always placed value on a landscape perspective.
The recent emergence of the discipline may be a rediscovery of the practical
value of a landscape perspective for understanding and managing nature.
The emergence of landscape ecology coincides with the widespread availability
of aerial photos (fig. 21.2) and satellite images (fig. 21.3). These
modem technologies offer overhead views of all landscapes, not just
those, such as Jomei's landscape around Mount Kagu,
that are conveniently located near heights.

FIGURE 21.2
Aerial photography has made the perspective sought by early Japanese
emperors accessible on all landscapes, such as this agricultural landscape
in

FIGURE 21.3
Images of the earth’s surface, such as this one of
Landscape ecology focuses on an organizational
scale above that addressed by community and ecosystem ecology. To a
landscape ecologist, a landscape
is a heterogeneous area composed of several ecosystems. The ecosystems
in a landscape generally form a mosaic of visually distinctive patches.
These patches are called landscape
elements. The elements in a mountain landscape typically include
forests, meadows, bogs, streams, ponds, and rock outcrops. An agricultural
landscape might include fields, fence lines, hedgerows, a
patch of woods, a farm yard, and dirt lane. An urban landscape usually
includes parks, industrial districts, residential areas, highways, and
sewage treatment works.
Landscape ecologists study landscape structure,
process, and change, In earlier chapters, we
discussed structure, process, and change in populations, communities,
and ecosystems. Structure, process, and change in landscapes form the
core of chapter 21.
CONCEPTS
l
Landscape structure includes
the size, shape, composition, number, and position of different ecosystems
within a landscape,
l
Landscape structure influences
processes such as the flow of energy, materials, and species between
the ecosystems within a landscape,
l
Landscapes are structured
and change in response to geological processes, climate, activities
of organisms, and fire.
CASE HISTORIES: landscape structure
Landscape structure includes the size, shape, composition,
number, and position of different ecosystems within a landscape.
Much of ecology
focuses on studies of structure and process; landscape ecology is no
exception. We are all familiar with the structure, or anatomy, of organisms.
In chapter 9 we discussed the structure of populations, and in chapters
16 to 20 we considered the structure of communities and ecosystems.
However, what constitutes landscape structure? Landscape
structure consists mainly
of the size, shape, composition, number, and position of ecosystems
within a landscape. As you look across a landscape you can usually recognize
its constituent ecosystems as distinctive patches, which might consist
of woods, fields, ponds, marshes, or towns. The patches within a landscape
form the mosaic that we call landscape structure
Most questions in landscape ecology require
that ecologists quantify landscape structure. The following examples
show how this has been done on some landscapes and how some aspects
of landscape structure are not obvious without quantification.
The Structure of Six Landscapes in
In 1981. G. Bowen and R. Burgess published a quantitative
analysis of several
First, let’s consider total forest cover.
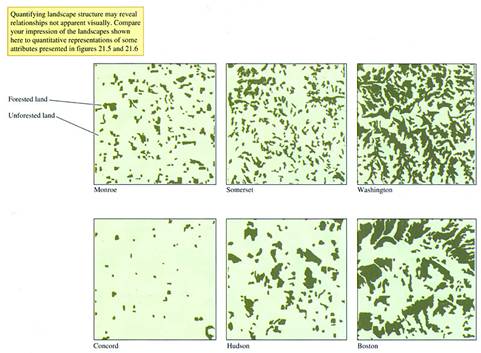
FIGURE 21.4
Forest fragments, shown as dark green, in six landscapes in
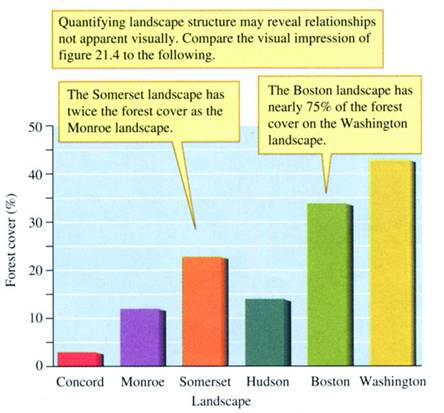
FIGURE 21.5 Percent
forest cover in six landscapes in
Now let's examine the size of forest patches
in each of the landscapes. Again, the median area of forest patches
differs significantly across the landscapes. The smallest median areas
are in the
Now. look back at figure
21.4 and try to estimate which of the landscapes contains the greatest
number, or highest density, of forest patches. The
Now let's look at a more subtle feature of
landscape structure, patch shape. Bowen and Burgess quantified patch
shape by the ratio of patch perimeter to the perimeter (circumference)
of a circle with an area equal to that of the patch. Their formula was:

where:
S =
patch shape
P =
patch perimeter
A =
patch area
How do you translate differences in the value
of this index into shape? If S is about equal to one, the patch is approximately
circular. Increasing values of S indicate less circular patch shapes.
High values of S generally indicate elongate patches and a long perimeter
relative to area.
Bowen and Burgess calculated the shapes, S,
for the forest patches in each of their landscapes and then determined
the median shape for each (fig. 21 .6). The

FIGURE 21.6 Relative shapes of forest patches
in six landscapes in
Until recently, geometry, which means "earth
measurement,'' could only offer rough approximations of complex landscape
structure. Today, an area of mathematics called fractal geometry can
be used to quantify the structure of complex natural shapes. Fractal
geometry was developed by Benoit Mandelbrot (1982) to provide a method
for describing the dimensions of natural objects as diverse as ferns,
snowflakes, and patches in a landscape. Fractal geometry offers unique
insights into the structure of nature.
The Fractal Geometry of Landscapes
During the
development of fractal geometry, Mandelbrot asked a deceptively simple
question: "How long is the coast of
Mandelbrot's answer to his question about
the British coastline was, "Coastline length depends on the scale
at which it is measured!" We can see the ecological significance
of this finding by considering some of its consequences to organisms.
Bruce Milne (1993) measured the coastline of Admiralty Island off the
coast of southeastern
Milne considered how the measured length of

FIGURE 21.7
Relationship between ruler length and the measured length of the coastline
of
Now, what "ruler" are bald eagles
and barnacles using? The distribution of eagle nests
around
Milne assumed that the eagles are, in effect,
using a ruler
spatial complexity around

FIGURE 21.8
Perspective on landscapes: fractal geometry tells us that the length
of coastline accessible to (a) the oil molecules spilling from
the hold of an oil tanker, such as the Exxon Valdez (the larger ship
shown in this photo) is much greater than that used by (b) bald eagles.
As in other areas of science, describing aspects
of landscape structure, such as the length of the coastline of Admiralty
Island or the size, shape, and number of forest patches in
CASE HISTORIES: landscape processes
Landscape structure influences processes such as the
flow of energy, materials, and species between the ecosystems within
a landscape.
Landscape
ecologists study how the size, shape, composition, number, and position
of ecosystems in the landscape affect landscape
processes. Though less familiar than physiological and ecosystem
processes, landscape processes are responsible for many important ecological
phenomena. In chapter 20, we saw how landscape structure, especially
the location of shallow bedrock, controls the exchange of nutrients
between subsurface and surface waters and local rates of primary production
in
Landscape Structure and the Dispersal of Small Mammals
Landscape
ecologists have proposed that landscape structure, especially the size,
number, and isolation of habitat patches, can influence the movement
of organisms between potentially suitable habitats. Populations of many
species occur in spatially isolated patches, with significant exchange
of individuals among patches. The group of subpopulations
living on such patches together make up a metapopulatlion. For instance, populations of desert bighorn
sheep live in the isolated mountain ranges of the southwestern

FIGURE 21.9
Fragmented landscapes: (a) the small isolated mountain ranges of the
southwestern United Slates and northern
Human activity often produces habitat fragmentation,
which occurs where a road cuts through a forest, a housing development
eliminates an area of shrubland, or tracts
of tropical rain forest are cut to plant pastures. Because habitat fragmentation
is increasing, ecologists study how landscape structure affects the
movements of organisms, movements that might mean the difference between
population persistence and local extinction.
James Diffendoffer,
Michael Gaines, and Robert Holt (1995) studied how patch size affects
the movements of three small mammal species: cotton rats, Sigmodon
hispidus, prairie voles, Microtus ochrogaster, and
deer mice, Peromyscus maniculatus. They divided
a
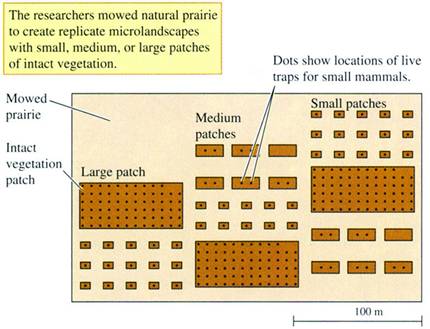
FIGURE 21.10
Experimental landscape for the study of small mammal movements
(data from Diffendorfer, Gaines, and Holt
1995).
The researchers predicted that animals would
move farther in the more fragmented landscapes consisting of small habitat
patches. In fragmented landscapes, individuals must move farther to
find mates, food, and cover. They also predicted that animals would
stay longer in the more isolated patches within fragmented landscapes.
Consequently, the proportion of animals moving would decrease with habitat
fragmentation.
The rodent populations were
monitored on the study site by trapping them with
The behavior of two of the three study species
supports the hypothesis that small mammals move farther in more fragmented
landscapes. As predicted, Peromyscus and Microtus, living in small patches, moved farther than individuals
living in medium or large patches (fig. 21.11). However the movements
of Sigmodon in medium and large patches did
not differ significantly.

FIGURE 21.11 Influence of patch size on small mammal
movements within experimental landscapes (data from Diffendorfer.
Gaines, and Holt 1995).
The proportion of Sigmodon,
Microtus, and Peromyscus
moving within the
In summary, this experiment shows a predictable
relationship between landscape structure and the movement of organisms
across landscapes. As the following example shows, those movements may
be crucial to maintaining local populations.
Habitat Patch Size and Isolation and the Density of Butterfly
Populations
Ilkka Hanski, Mikko Kuussaari, and Marko Nieminen (1994) found that the local population density of
the Glanville fritillary butterfly, Melitaea
cinxia, is significantly affected by the size and isolation
of habitat patches. The researchers studied a metapopulation
of these butterflies on

FIGURE 21.12
Much of the landscape of southwestern
There were 50 patches of potential habitat
within the study area. Forty-two of these patches were occupied by the
butterflies in 1991. The patches ranged in area from 12 to
Habitat patch area influenced both the size
and density of the populations. Total population size within a patch
increased with patch area. However, population density decreased as
patch area increased (fig. 21.13). Thus, though large habitat patches
supported larger numbers of individuals than smaller patches, population
density was lower on large patches.

FIGURE 21.13
Relationship between habitat patch area and population size and density
of the butterfly Melitaea cinxia
in a landscape on
The team also found that more isolated patches
supported lower densities of butterflies. Isolation influences local
population density in these populations because local populations are
partly maintained by immigration of Melitaea
from other patches. For instance, during I
week of sampling the butterflies in one patch, about 15% of the males
and 30% of the females were recaptures from surrounding patches.
This experiment determined that area and isolation
of patches strongly influence the size and density of Melitaea populations. One conclusion that we can draw from
these patterns is that landscape structure is important for understanding
the distribution and abundance of the butterflies. It turns out that
landscape structure also affects the persistence of local populations.
Between 1991 and 1992, Hanski and his colleagues
recorded three extinctions of local populations and five colonizations
of new habitat patches. All these extinctions and colonizations
occurred in small patches with small populations.
The vulnerability of small populations to
extinction has been well documented in populations of desert bighorn
sheep in the southwestern
The studies by Diffendorfer
and colleagues, Hanski and colleagues, and
Berger show that the movement of organisms and the characteristics of
local populations are significantly influenced by landscape structure.
As we will see in the next example, landscape structure can also influence
the characteristics of ecosystems.
Landscape Position and
Katherine
Webster and her colleagues (1996) at the Center for Limnology at the
Webster and her colleagues set out to determine
whether the contrasting chemical responses of lakes to drought can be
explained by the position of the lake in the landscape. They worked
in northern
The sources of water for a lake are precipitation,
surface water, and groundwater flow. Different lakes receive different
proportions of their water from these sources, and these proportions
depend upon a lake's position in the landscape. Figure 21.14 shows a
series of lakes along a hydrologic flow system in northern

FIGURE 21.14
The important point here is that the positions
of these lakes in the landscape determine the proportion of water they
receive as groundwater. Webster and her colleagues estimated that
The responses of these seven lakes to a drought
were studied from 1986 to 1990. As you might expect, the levels of the
lakes dropped during this 4-year drought. However, the amount of drop
in lake level was related to a lake's position in the landscape (fig.
21.15). The level of

FIGURE 21.15
Landscape position also significantly influenced
a lake's chemical responses to the drought. The concentrations of dissolved
ions such as calcium (Ca2+) and magnesium (Mg2+)
increased in the majority of the lakes. However, the increase in ion
concentration was highest at the upper and lower ends of the hydrologic
flow system. Meanwhile, the combined mass of Ca2+ and Mg2+
increased in the three lakes at the lower end of the hydrologic flow
system but did not change in
The researchers concluded that the increased
mass of Ca2+ and Mg2+ seen at the lower end of
the hydrologic flow system was due to an increased proportion of inflows
from groundwater and surface water, sources rich in Ca2+
and Mg2+. The declines in mass of Ca2+ and Mg2+ in
In the first section of this chapter, we reviewed
the concept of landscape structure. In this section, we explored the
connection between landscape structure and landscape processes. But
what creates landscape structure? Landscape structure,
like the structure of populations, communities, and ecosystems, changes
in response to an interplay between dynamic processes. We explore
the sources of landscape structure and change in the next Case Histories
section.
CASE HISTORIES: origins of landscape structure and change
Landscapes are structured and change in response to geological
processes, climate, activities of organisms, and fire.
What creates
the patchiness we see in landscapes? Many forces combine in numerous
ways to produce the patchiness that we call landscape structure. In
this section, we review examples of how geological processes, climate,
organisms, and fire contribute to landscape structure.
Geological Processes, Climate, and Landscape Structure
The geological
features produced by processes such as volcanism, sedimentation, and
erosion provide a primary source of landscape structure. For instance,
the alluvial deposits along a river valley provide growing conditions
different from those on thin, well-drained soils on nearby hills. A
volcanic cinder cone in the middle of a sandy plain offers different
environmental conditions than the surrounding plain (fig. 21.16). Distinctive
ecosystems may develop on each of these geological surfaces, creating
patchiness in the landscape. In the following example, we shall see
how distinctive soils contribute to vegetative patchiness in a

FIGURE 21.16 Geological
features such as the volcanic cinder cone in the middle of a sandy plain
add structure to the landscape by adding a geological surface with distinctive
physical and chemical properties.
Soil and Vegetation Mosaics in the
The
From a distance, the bajadas
of the
McAuliffe established study sites on the bajadas of three mountain ranges. At each site he studied
soils and plant distributions. In all three study areas, he found a
wide range of soil types and plant distributions that corresponded closely
to soil age and structure.
Let's look at some of the patterns McAuliffe
found on the bajada associated with the northern
end of the
the bajada, to the
right in figure 21.17, the next soils in the sequence date from the
middle to late Pleistocene and are hundreds of thousands of years old.
These soils are followed by Holocene deposits that are less than 11,000
years old and are associated with an ephemeral desert water course called
Wildhorse Wash. Near
the Holocene soils. McAuliffe found soils that dated from the late Pleistocene.
These soils were 25,000 to 75,000 years old.

FIGURE 21.17
Soil ages on an outwash plain, or bajada,
associated with the
In the space of a few kilometers, McAuliffe
found patches of soil that were (I) almost 2 million years old, (2)
hundreds of thousands of years old, (3) tens of thousands of years old,
and (4) less than 11,000 years old. Because soil-building processes
occur over long periods of time, these soils of vastly different ages
also differ substantially in structure. Figure 21.18 shows McAuliffe's
drawings of typical profiles of Holocene, middle to late Pleistocene.
and early Pleistocene soils. The Holocene soils
had low amounts of clay and calcium carbonate (CaCO3) and
poorly developed soil horizons. They also lacked a caliche layer, a hardpan soil horizon formed
by precipitation of CaCO3. Middle to late Pleistocene soils
bad a much higher clay content than Holocene soils, and early Pleistocene
soils contained even more clay. These clay layers in the older soil
profiles are called argyllie horizons. Middle to
late and early Pleistocene soils also contained more CaCO3
and were underlain by a thick layer of caliche.
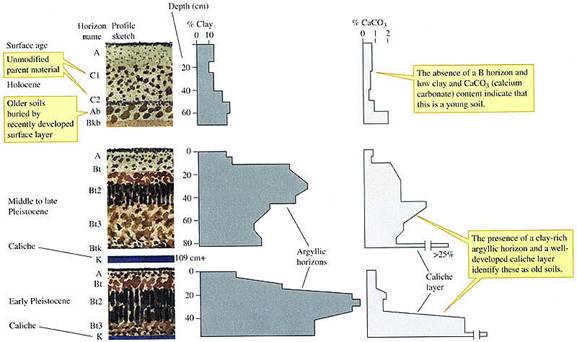
FIGURE 21.18
Structural features of young to old desert soils on the
These differences in soil structure influence
the distributions of perennial plants across the

FIGURE 21.19 Association
between vegetation and soils of different ages and structure on the
Climate and Landscape Structure
Climate is
a major determinant of landscape structure. Our review of major terrestrial
environments in chapter 2 showed a clear connection between landscape
structure and climate. Climate determines whether the potential ecosystem
in an area will be tundra, temperate forest, or desert. It also sets
the baseline for aquatic ecosystems. Climate determines whether rivers
flood once a year, twice a year, or at irregular intervals. As climate
changes, landscapes change. The advances and retreats of glaciers have
shaped whole continents, creating lakes and plains, transporting soils,
and carving mountain valleys. Wetting and drying cycles have changed
the distribution extent of rain forest and savanna in the
We can also see the signature of climate on
a very local scale. Let's go back to the soils studied by McAuliffe
and review some of the effects of climate. The soil mosaic along the
bajada east of the
These alluvial deposits were gradually changed,
and these changes were dependent upon climate. Two of the prominent
features of the older soils studied by McAuliffe were the formation
of a clay-rich argyllic horizon and the formation
of a CaCO3-rich caliche layer.
Both these soil features are the result of water transport. Clay particles
are transported as a colloidal suspension, while the CaCO3
is transported in dissolved form. Consequently, the clays precipitate
out of suspension higher in the soil profile than the CaCO3.
The result is the layering of an argyllic
horizon over a caliche layer as shown in figure
21.18.
Water, working on alluvial deposits, is responsible
for the soil structure observed by McAuliffe, but it was water delivered
to the landscape under particular climatic conditions. We can get a
clue about those conditions by observing some soil characteristics.
We know that argyllic horizons are deposited by water. However, the soils
described by McAuliffe also offer clues that the action of water was
highly episodic. The argyllic horizon in these
soils is red, and this red color is the result of a buildup of iron
oxides. Oxidation of iron could have only occurred in an oxidizing environment.
Because soil saturated with water quickly becomes anoxic, the presence
of oxidized iron in the argyllic horizon indicates
that these soils were formed when conditions were intermittently wet.
In other words, the soils along the bajada
of the
While geological processes and climate set
the basic template for landscape structure, the activities of organisms
can be an additional source of landscape structure and change. In the
following section, we consider how the activities of humans and other
species can change landscape structure.
Organisms and Landscape Structure
Organisms
of all sorts influence the structure of landscapes. While the following
discussion focuses on the influences of animals, plants create much
of the distinctive patchwork we call landscape structure. For an example
of how plants can change landscape structure, think back to chapter
19, where we discussed Edward Witkowski's studies (1991) of how Acacia affects the South
African fynbos. In that discussion, we focused
on the effects of the plant on the quantity of soil nitrogen and rates
of decomposition. Now let's take a landscape perspective of the effects
of Acacia. As this plant invaded the fynbos,
it created distinctive patches where the availability of nitrogen is
higher and where decomposition rates are higher. By adding these distinctive
ecosystem patches, Acacia has altered landscape structure.
Many studies of landscape change have focused
on the conversion of forest to agricultural landscapes. In

FIGURE 21.20
Human-caused change in forest cover in
In eastern
Figure 21.21 shows the changes in the composition
of the Veluwe landscape from 1845 to 1982.
The greatest change over this period was a shift in dominance from heathlands to forests. In 1845, heathlands
made up 66% of the landscape, while forests constituted 17%. By 1982,
coverage by heathlands had fallen to 12% of
the landscape and forest coverage had risen to 64%. The figure also
shows modest but ecologically significant changes in the other landscape
elements. The area of drift sand reached a peak in 1898 and then dropped
and held steady at 3% to 4% from 1957 to 1982. Urban areas established
a significant presence beginning in 1957. Finally. coverage by agricultural
areas has varied from 9% to 16% over the study interval, the least variation
shown by any of the landscape elements.

FIGURE 21.21 Change in a Dutch landscape
(data from Hulshoff 1995).
As total coverage by forest and beathlands changed within the Veluwe
landscape, the number and average area of forest and heath patches also
changed. These changes indicate increasing fragmentation of heathlands
and decreasing fragmentation of forests. For instance, between 1845
and 1982, the number of forest patches declined, while the average area
of forest patches increased. During the same period, the number of heath
patches increased until 1957. Between 1957 and 1982, the number of heath
patches decreased as some patches were eliminated entirely. The average
area of heath patches decreased rapidly between 1845 and 1931 and then
remained approximately stable from 1931 to 1982.
During the period that
As we enter the twenty-first century, economically
motivated human activity continues to change the structure of landscapes
all over the globe. We examine current trends in land cover at the global
scale in chapter 23. Before we do that, however, let's examine the effects
of some other species on landscape structure.
Many animal species modify landscape structure
(fig. 21.22). African elephants feed on trees and often knock them down
in the process. As a consequence, these elephants can gradually change
woodland m grassland. Alligators maintain ponds in the Florida Everglades,
a landscape element upon which many species depend to survive droughts.
Small species can also change landscapes. Kangaroo rats, Dipodomys
spp., of the American Southwest dig burrow systems that modify
the structure of the soil, the distribution of nutrients, and the distribution
of plants to such an extent that the result is recognizable from aerial
photos. Similar effects on landscape structure are created by termites
and ants.

FIGURE 21.22
Species with significant impacts on landscape structure. (a) African
elephants control the extent of tree cover in some landscapes. (b) Alligators
build and maintain ponds in wetland landscapes. (c) Feeding and burrowing
by kangaroo rats introduce added patchiness into desert landscapes.
(d) Termite mounds add distinctive landscape features.
One of the most adept modifiers of landscapes
is the beaver, Castor canadensis (fig. 21.23).
Beavers alter landscapes by cutting trees, building dams on stream channels,
and flooding the surrounding landscape. Beaver dams increase the extent
of wetlands in the landscape, alter the hydrologic regime of the catchment, and trap sediments, organic matter, and nutrients.
The selective cutting of trees adds patchiness to the plant community
and reduces the abundance of tree species preferred as food. These effects
add several novel ecosystems to the landscape.

FIGURE 21.23 Beavers are among nature's
most active landscape engineers.
These influences of beavers on landscape structure
once shaped the face of entire continents. At one time, beavers modified
nearly ail the temperate stream valleys in the Northern Hemisphere.
The range of beavers in North America extended from arctic tundra to
the Chihuahuan and
Carol Johnston and Robert Naiman and their colleagues have carefully documented the
substantial effects of beavers on landscape stmcturu
(e.g., Naiman et al. 1994). Much of their
work has focused on the effects of beavers on the
Beaver activity has changed the
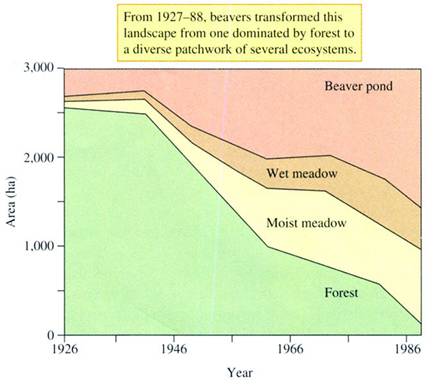
FIGURE 21.24
Beaver-caused landscape changes on the
Naiman and his colleagues quantified the effects
of beaver over
The changes in landscape structure
induced by beavers substantially alter landscape processes such as nutrient
retention. Beaver activity between 1927 and 1988 increased the quantity
of most major ions and nutrients in the areas affected by impoundments
(fig. 21.25). The total quantity of nitrogen increased
by 72%, while the amounts of phosphorus and potassium increased by 43%
and 20%, respectively. The quantities of calcium, magnesium,
iron, and sulfate stored in the landscape were increased by even greater
amounts.

FIGURE
21.25 Nutrient retention
on the
Naiman and his colleagues offer three possible explanations
for increased ion and nutrient storage in this landscape: (1) beaver
ponds and their associated meadows may trap materials eroding from the
surrounding landscape, (2) the rising waters of the beaver ponds may
have captured nutrients formerly held in forest vegetation, and (3)
the habitats created by beavers may have altered biogeochemical processes
in a way that promotes nutrient retention. Whatever the precise mechanisms,
beaver activity has substantially altered landscape structure and processes
on the
Fire and the Structure of a Mediterranean Landscape
Fire contributes
to the structure of landscapes ranging from tropical savanna to boreal
forest. However, fire plays a particularly prominent role in regions
with a Mediterranean climate. As we saw in chapter 2, terrestrial ecosystems
in regions with Mediterranean climates, which support temperate woodlands
and shrublands, are subject to frequent burning. Hot, dry summers
combined with vegetation rich in essential oils create ideal conditions
for fires, which can be easily ignited by lightning or humans. In regions
with a Mediterranean climate, fire is responsible for a great deal of
landscape structure and change.
Richard Minnich
(1983) used satellite photos to reconstruct the fire history of southern
Minnich proposed that the different fire histories
of southern
Minnich tested his hypothesis using satellite images
taken from 1972 to 1980 (fig. 21.26). He found that between 1972 and
1980 the total area burned in the two regions was fairly similar (fig.
21.27). However, the size of burns differed significantly between the
two regions. The frequency of small bums below

FIGURE 21.26
Areas of temperate shrubland in southern
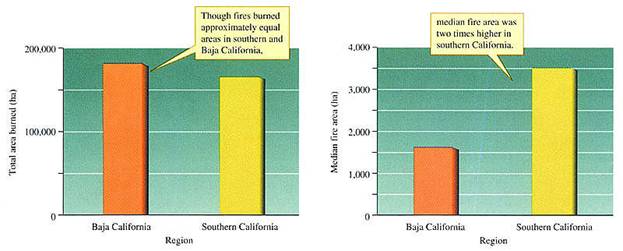
FIGURE 21.27
Characteristics of fires in the Mediterranean landscapes of southern
and
These results are consistent with Minnich's hypothesis, but do they show conclusively that differences
in fire management in southern
In this section, we have seen how geological
processes, climate, the activities of organisms, and fire can contribute
to landscape structure and change. Because human activity has often
greatly altered landscape structure, there is growing interest in landscape
restoration. That is the subject that we take up in the Applications
and Tools section.
APPLICATIONS AND TOOLS: restoring a riverine landscape
Rivers and
their floodplains form a complex, highly dynamic landscape that includes
river, riparian forest, marsh, oxbow lake, and wet meadow ecosystems.
Historically, these ecosystems actively exchanged organisms, inorganic
nutrients, and organic energy sources. The key linkage between these
landscape elements was periodic flooding.
Floods connect rivers with their associated
floodplain ecosystems, and the rates and timing of many ecological processes
are determined by the "flood-pulse" (Junk, Bayley,
and Sparks 1989, Bayley 1995). Floods determine
the form of the riverine landscape by depositing
silt on floodplains, isolating oxbow lakes, and creating new river channels.
Floods increase rates of decomposition and nutrient cycling in floodplain
environments. Many species of river fish use floodplains as spawning
and nursery grounds and many riparian plants require flooding for germination
and establishment.
Over the past century, water management by
building dams, channelizing rivers, constructing
flood levees, and diverting water for irrigation has cut the historic
connections between most rivers and their floodplains. However, there
is growing recognition of the value of these historic connections for
maintaining water quality and for supporting biological diversity. Consequently,
governments all over the world have begun programs of river restoration.
Some of the most ambitious of these projects focus on the
Riverine
Restoration: The
The Kissimmee
River flows from its headwaters in
Before flood control, the relationship of
the Kissimmee River to its floodplain was similar to that of large tropical
rivers such as the Amazon River in South America and Niger River in
Rapid human population growth in the early
1940s, followed by extensive flooding that lasted from 1947 to 1948,
created pressures for flood control on the

FIGURE 21.28 Channelization and wetland loss in the

FIGURE 21.29
Channelizing great(v simplified the
structure of the
These environmental changes had a severe impact
on many populations and ecosystem processes.
Wintering water-fowl populations declined by 92%. The population of
large-mouth bass, an important sport fish, declined. Largemouth bass
were replaced by nongame species that tolerate low oxygen concentrations. Populations
of riverine invertebrates were reduced and
replaced by invertebrates of lake and pond ecosystems. Eliminating the
flood pulse greatly reduced the exchange of nutrients, organic matter,
and organisms between the river and floodplain ecosystems. Stabilized
water levels nearly eliminated spawning and foraging habitats for adult
fish and refuge and rearing areas for larval and juvenile fish. Ecologists
estimated that drying the floodplain wetlands along the
The public soon recognized the negative ecological
consequences of the
The
In response to these encouraging results,
water managers began a far more ambitious plan to restore a large portion
of the

FIGURE 21.30
Restoration of the
The keys to this restoration effort involve
restoration of landscape structure and landscape processes. First, water
managers will restore historical landscape structure by restoring the
complex channel network and reestablishing the historic floodplain marshes.
Second. they will restore the dominant landscape process, the historic
flood pulse. Restoration of flooding will reestablish the hydrologic
connections between the river and floodplain ecosystems, connections
that promote exchanges of nutrients, energy, and species among ecosystems
in the
This is the largest landscape restoration
project ever undertaken. The restoration of the
SUMMARY CONCEPTS
A landscape
is a heterogeneous area composed of several ecosystems. The ecosystems
making up a landscape generally form a mosaic of visually distinctive
patches. These patches are called landscape clements.
Landscape ecology is the study of landscape structure and processes.
Landscape
structure includes the size, shape, composition, number, and position
of different ecosystems within a landscape. Most questions in landscape ecology require that ecologists quantify landscape
structure. Until recently,
however, geometry, which means “earth measurement,” could offer only
rough approximations of complex landscape structure. Today, an area
of mathematics called fractal geometry can be used to quantify the structure
of complex natural shapes. One of the findings of fractal geometry is
that the length of the perimeter of complex shapes depends upon the
size of the device used to measure the perimeter. One implication of
this result is that organisms of different sizes may use the environment
in very different ways.
Landscape
structure influences processes such as the flow of energy, materials,
and species between the ecosystems within a landscape. Landscape ecologists have proposed that landscape structure,
especially the size, number,
and isolation of habitat patches, can influence the movement of organisms between potentially suitable habitats. The group of subpopulations living on such habitat patches make
up a metapopulation.
Studies of the movements of small mammals
in a prairie landscape show that a smaller proportion of individuals
moves in more fragmented landscape but that the individuals that do move will move farther.
The local population density of the Glanville fritillary butterfly,
Melitaea cinxia, is lower on larger and on
isolated habitat patches. Small
populations of this butterfly and desert bighorn sheep are more vulnerable to local extinction. The source of water for lakes in a
Landscapes
are structured and change in response to geological processes, climate,
activities of organisms, and fire. Geological features produced by processes such as volcanism,
sedimentation, and erosion interact with climate to provide a primary source of landscape structure. In the
Because human activity has often altered landscape
structure and processes in undesirable ways, there is growing pressure
and interest in landscape restoration. Some of the most ambitious current
restoration efforts focus on the restoration of riverine
landscapes. Rivers and their floodplains form a complex, highly dynamic
landscape that includes river riparian forest, marsh, oxbow lake, and
wet meadow' ecosystems. Over the past century, water management by building
dams, channelizing rivers, constructing flood levees, and diverting
water for irrigation has cut the historic connections between most rivers
and their floodplains. The restoration of the
REVIEW
QUESTIONS
1. How does landscape ecology
differ from ecosystem and community ecology? What questions might an
ecosystem ecologist ask about a forest? What questions might a community
ecologist ask about the same forest? Now. what
kinds of questions would a landscape ecologist ask about a forested
landscape?
2. How should the area of forest
patches in an agricultural landscape affect the proportion of bird species
in a community that are associated with forest edge habitats? How should
patch area affect the presence of birds associated with forest interiors?
3. The green areas represent forest
fragments surrounded by agriculture. Landscapes I and 2 contain the
same total forest area. Will landscape I or 2 contain more forest interior
species? Explain.
4. How might the shapes of forest
patches in a landscape affect the proportion of birds in the community
associated with forest edge habitat? How might patch shape affect the
presence of birds associated with forest interior?
5. Consider the following options
for preserving patches of riverside forest. Again, the two landscapes
contain the same total area of forest but the patches in the two landscapes
differ in shape. Which of the two would be most dominated by forest
edge species?
6. How does the concept of metapopulations differ from the perspective of populations
that we discussed in section III? (Hint: Think of the spatial contexts
of these two views of populations.)
7. How do the positions of patches
in a landscape affect the movement of individuals among habitat patches
and among portions of a metapopulation? Again. consider the hypothetical landscapes
shown in question 5. Which of the two landscapes would promote the highest
rate of movement of individuals between forest patches? Can you think
of any circumstances in which it might be desirable to reduce the movement
of individuals across a landscape? (Hint: Think of the potential threat
of pathogens that are spread mainly by direct contact between individuals
within a population.)
8. Use fractal geometry and the
niche concept (see chapters 13 and 16) to explain why the canopy of
a forest should accommodate more species of predaceous insects than
insectivorous birds. Assume that the numbers of bird and predaceous
insect species are limited by competition. (Milne's study [1993] of
barnacles and bald eagles on
9. Analyses such as Milne's comparison
(1993) of bald eagles and barnacles demonstrate that organisms of different
sizes interact with the environment at very different spatial scales.
With this in mind consider the experiments of Diffendorfer
and colleagues (1995) on the influence of habitat fragmentation on movement
patterns of small mammals. Think about the size of their experimental
study area (see fig. 21.10). How might a manipulation of this size have
affected the movements of prairie birds? How would their manipulation
have affected the movements of ground-dwelling beetles?
10. How do the activities of animals
affect landscape heterogeneity? You might use either beaver or human
activity as your model. What parallels can you think of between the
influence of animal activity on landscape heterogeneity and the intermediate
disturbance hypothesis? Which is concerned with the effect of disturbance
on species diversity?